Male-male competition has never been contested and easy to understand. Associated with sexual dimorphism in many species. Need to be careful, however, as there are other causes for sexual dimorphism. Cardinals are monogamous and the bright red color of the male is attributed to his acting as a decoy for predators to protect his family.
Female choice was controversial and initially defenders of Darwin's theory such as Huxley, doubted it was of any influence in shaping mating systems.
One of the problems is determining what a female should be looking for in a mate?
Species recognition
Resources
"Good genes"
Most evolutionists would jump on good genes as the criteria females should be selection, but then what are good genes?
In the beginning, evolutionists simply proposed it was sexual attractiveness or the sexy son hypothesis.
"In a society where males compete with each other to be chosen as he-men by females, one of the best things a mother can do for her genes is to make a son who will turn out in his turn to be an attractive he-man. If she can ensure that her son is one of the fortunate few males who wins most of the copulations in the society when he grows up, she will have an enormous number of grandchildren. The result of this is that one of the most desirable qualities a male can have in the eyes of a female is, quite simply, sexual attractiveness itself" Richard Dawkins, The selfish gene, 1976.
But this begs the question of why a tail should be attractive in the first place, especially since you can demonstrate survival "costs" to this trait. Answer question one.
Three of the most cited theories why exaggerated traits such as a peacock tail are sexually attractive:
One: an old theory (R. A. Fisher, The Genetical Theory of Natural Selection, Clarendon Press, Oxford, 1930).
"Runaway selection" on ornaments ": Females prefer males with certain characteristics, so immediately not only are males who have the trait favoured but the preference itself. This is bound to lead to the evolution of preferences for more exaggerated traits and more pronounced preferences. If females prefer suddenly a red spot, then not only are males with a red spot favoured but females who prefer red spots and through time, males with larger red spots and females who prefer larger red spots. The limit to this process would be considerations of survival and once this limit was met, the trait should over time become fixed in the population.
Two: A new theory, the Handicap principle (A. Zahavi. 1975. J. Theor. Biol, 53: 205-214). Organisms are honest signalers of quality because the signals used are costly and so can only be used or displayed in individuals of good quality. So a female prefers a male with a handicap because this male must have high viability (good genes), to be able to survive with the handicap
"An individual with a well developed sexually selected character [such as a peacock's flashy tail] is an individual which has survived a test. A female which could discriminate between a male possessing a sexually selected character, from one without it, can discriminate between a male which has passed a test and one which has not been tested. Females which selected males with the most developed characters can be sure that they have selected from among the best genotypes of the male population. " Amotz Zahavi (1975)
Theory Three: Parasite mediated sexual selection. Skim W. D. Hamilton, M. Zuk. 1982. Science. Vol 218, Issue 4570, 384-387.
Traits that are exaggerated are those that indicate a males fitness with regard to parasites and pathogens. First proposed as a special case of the handicap principle.They found a highly significant association in species of North American birds between incidence of chronic blood infections and striking display characters.
Essentially we have two basic theories with one good general case study for the second. Both theories. Fisher's and Zahavi's, incorporate serious theoretical problems. The problem with both theories is the connection between good genes and the character in question.
Fisher explains maintenance but not how such a preference becomes established in the first place. So all sorts of questions on the nature of the genetic bases of such preferences and on how "linked" do preference have to be with fitness to become established. What also should the deferential survival among males be to get runaway selection established with regard to preferences, since the model indicates females initially choose on a relative scale. Note however, that once runaway selection is taking place, only large decreases in relative survivorship among males with and without exaggerated traits will stop the continual exaggeration for the trait.
Fisher later in his text acknowledges a possible connection between quality and the character at least initially (not needed later in the runaway process), but really says nothing about how the connection evolved. Examine the diagram below for some ideas on how a trait could be exaggerated using Fisher's ideas
.
Zahavi's theory does not suffer as much from this criticism, since all you need is some handicap and an average advantage for honesty, but still suffers from lack of discussion regarding how the correlation between quality and the trait is maintained. Zahavi's theory needs males to be honest salesmen. Problem is that environmental change can change the handicap and there can be recombination between fitness and handicap. Therefore it is possible to get progeny of lower quality sporting the same handicap as males of high quality.
Example:
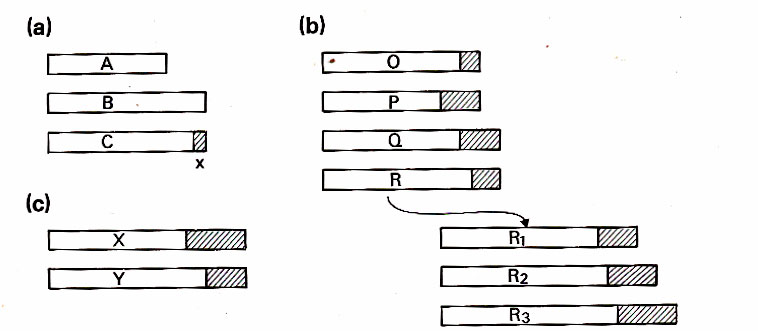
(a) Basic choice in Zahavi's scheme: Males A and B differ in quality but are identical in appearance and thus cannot be differentiated by females. Male C is as good as B but carries a marker which constitutes a handicap, reducing his quality by an amount X. Females choosing C will pass on more quality to the sons than those choosing A, but less than those choosing B. However since females choosing B are equally likely to choose A, they will as a type, on average, pass on less to their sons than those choosing C, So there is selection for C-type-male-choosing females in the population over time.
(B) Problems with trait exaggeration in Zahavi's model.
P males will not be selected out of population, even though they are of lower quality than R males. They are in an evolutionary sense "dishonest" salesme.
Q and O are equal so there is no advantage for females in terms of reproductive fitness to choose Q. Yet females choosing on the basis of an handicap should choose both P and Q. Females should be choosing R or highest quality males. But they do not if choosing on the bases of the handicap because now P and Q males are "dishonest " salesmen.
Increased reproduction because of quality goes to the females who choose R, but any increase in trait has to be correlated with a change in overall quality. The problem is for the trait to be come very exaggerated, quality also has to increase. There’s got to be a limit to quality.
(c) Not addressed initially by Zahavi, is any positive change in the handicap(X-Y) which is costly, can cause a decrease in male quality and makes males dishonest salesmen relative to other males. So any change in the amount of the handicap displayed has to be strictly correlated with a relative change in quality.
The only way at this stage an handicap can increase is if it is an indicator of male fitness but not strictly genetically fixed. A male who can survives despite a broken leg might be demonstrating to females that he can survive despite this handicap and in reality this may be true, but only his quality not his handicap will be passed on. So females may start looking for injured males who survived. Although this example might seem farfetched, think about Zuk and Hamilton's work in which parasite resistance is passed down, but not necessary the parasite infection itself. But females appear to gain at least comparatively (interspecific comparisons between species subject and not as subject to parasite infection) in species where parasites are common to mate with males who can still demonstrate bright plumage and costly displays.
Here is one real life non-parasite/ pathogen example that would fit Zahavi's theory and ensure honest salesmanship or a continuing connection between trait exaggeration and male quality.
This is a trait that is inherited and refers to crests that develop only in breeding season. However, the extent to which a male can develop the crest will depend on whether he can sequester the resources to do it. So crest size is not genetically fixed, is an indicator of male superiority and there is selection to mate with large crested individuals. Example: Trilurus newts.
Male in breeding condition on your left.
Females are grasped by male. Male’s ability to sustain courtship is important and related to his capacity to hold his breath. His crest being vascularized may act as an additional respiratory surface. In an experiment newts deprived of access to the atmosphere survive for longer if they are in breeding condition.
Using the information in this guide and Hamilton and Zuk's paper, answer questions 2-5. Turn in answers to questions 1-5 for homework.